New paper (behind paywall) Sahelian pastoralism from the perspective of variants associated with lactase persistence, by Priehodová et al. Am J Phys Anthropol (2020) e24116.
Interesting excerpts from the discussion (emphasis mine, minor modifications for clarity):
Our investigation of LP variant frequencies revealed new and interesting results related to the origins of pastoralism and subsequent gene flow between pastoralists and farmers in the Sahel/Savannah belt of Africa.
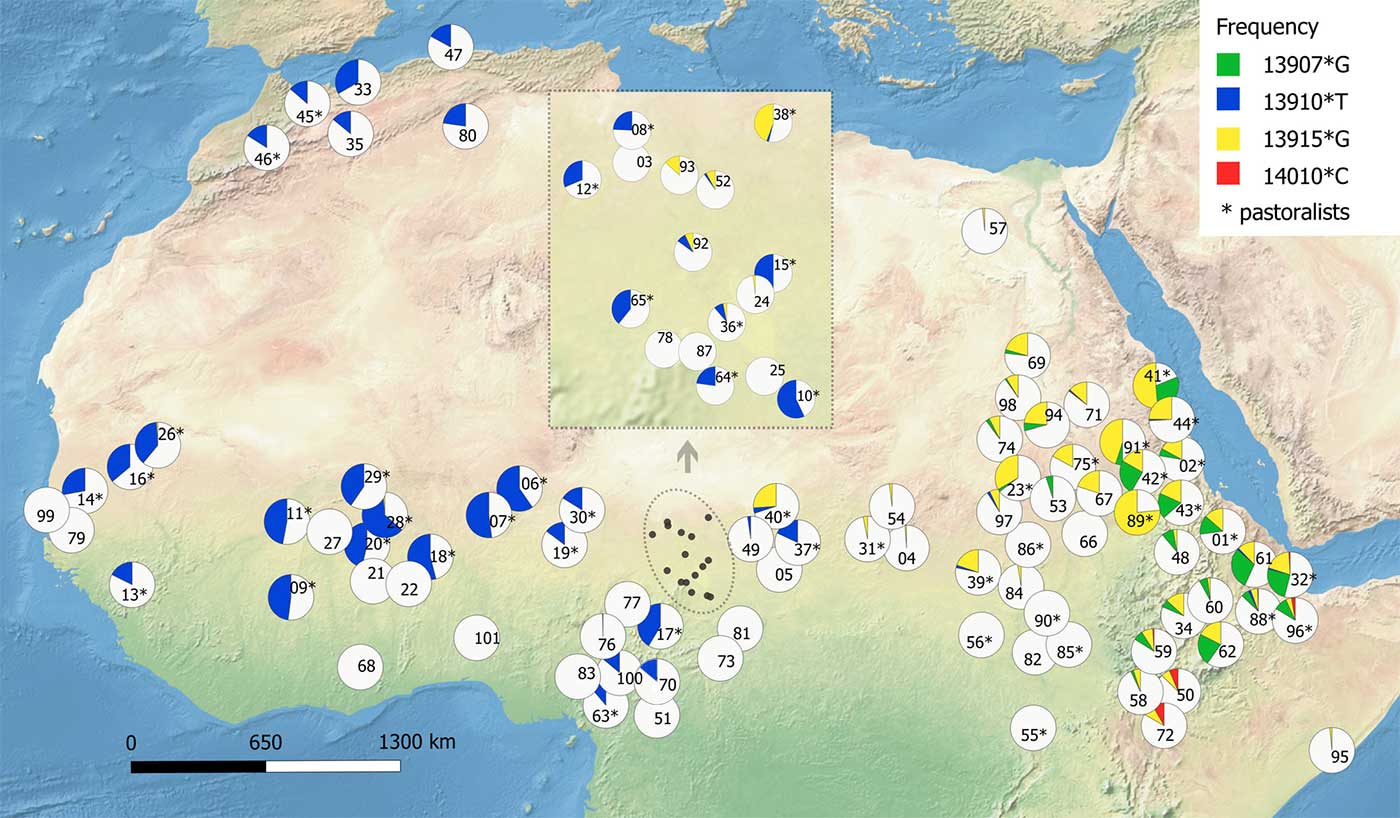
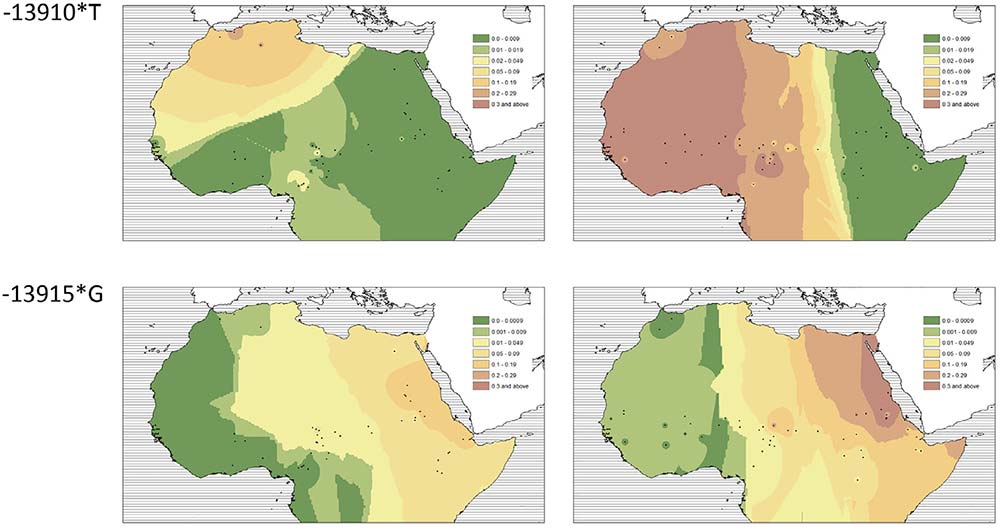
- We observed a clear distinction between regions west and east of Lake Chad: while variant −13910*T prevails in the western Sahel, where we found it only in pastoralists such as the Fulani, Tuareg, and Moors (it is virtually absent in sedentary farmers analyzed in our dataset), eastern Sahel appears to be much more diverse and several LP-associated variants, such as −13907*G, −13915*G, and 14010*C, are found in various populations.
- The almost zero frequency of the −13910*T variant in sedentary farmers is consistent with a hypothesis of very low gene flow in the western part of the Sahel/Savannah belt. Low gene flow in this region is further attested by analyses of mtDNA variation (Čížková et al., 2017; Kulichová et al., 2017) and supported by recent geometric morphometrics analyses (Kleisner, Pokorný, Čížková, Froment, & Černý, 2019).
- The presence of the “Arabian” −13,915*G variant in many populations of the eastern Sahel (including several sedentary farmers) is, on the other hand, consistent with the idea of a higher gene flow in this part of Africa (Čížková et al., 2017), especially between the Arabic-speaking Baggara and their neighbors (Priehodová et al., 2017).
- The −14009*G variant has been associated with LP in eastern Africa (Jones et al., 2013; Jones et al., 2015) and frequency at which we found it in the Beja near Kassala, Sudan (42.6%) is much higher than reported by other studies (Hassan et al., 2016; Ranciaro et al., 2014). Such isolated occurrence and high frequency of this mutation among the Beja specifically could be explained by East African origin of this Cushitic-speaking population and subsequent genetic drift with a limited gene flow, which has been attested also by an mtDNA study (Čížková et al., 2017).
- Interestingly, in all Nilotic pastoralists analyzed in this study we found zero frequencies of the main known LP variants. It is possible that these populations also carry another, yet unknown, LP variant since tests for lactose tolerance (…)
Our study shows that the age of expansion of −13910*T obtained for the Fulani pastoralists in western Sahel/Savannah belt (7,534–9,686 years ago) is much older than that obtained for the −13915*G in Arabic pastoralists in eastern Sahel/Savannah belt (Priehodová et al., 2017), when applying the same method (Austerlitzet al., 2003). Priehodová et al., 2017 estimated age of −13915*G at between 1,274 and 1,782 years ago, which is consistent with the time of Arabic expansion to Africa. The estimate obtained in this study for the Fulani pastoralists conforms to estimates for introduction of pastoralism in Africa which are based on archeological evidence. There is clear evidence of early Middle Holocene migration(s) of pastoralists from the Near East to northeast Africa (Jesse et al., 2013; Kuper & Riemer, 2013) as well as of the fact that these pastoralists later established themselves in western Africa (Linseele, 2013; Ozainne et al., 2014). It has been suggested that presence of the −13910*T variant in North African Berber population could be a sign of past gene flow from Near Eastern pastoralists (Myles et al., 2005).
This first Saharan/Sahelian influx of population with LP individuals was followed by a later migration of other pastoralists from the Arabian Peninsula throughout northern Africa and Sahara up to the Lake Chad Basin, which was led by Bedouin Arabic-speaking tribes (Braukämper, 1994; Levy & Holl, 2002; Zeltner, 2002). This migration probably introduced the −13915*G variant to the region, which originated in Arabia as an adaptation to the consumption of camel milk (Enattah et al., 2008; Imtiaz et al., 2007; Priehodová et al., 2014).
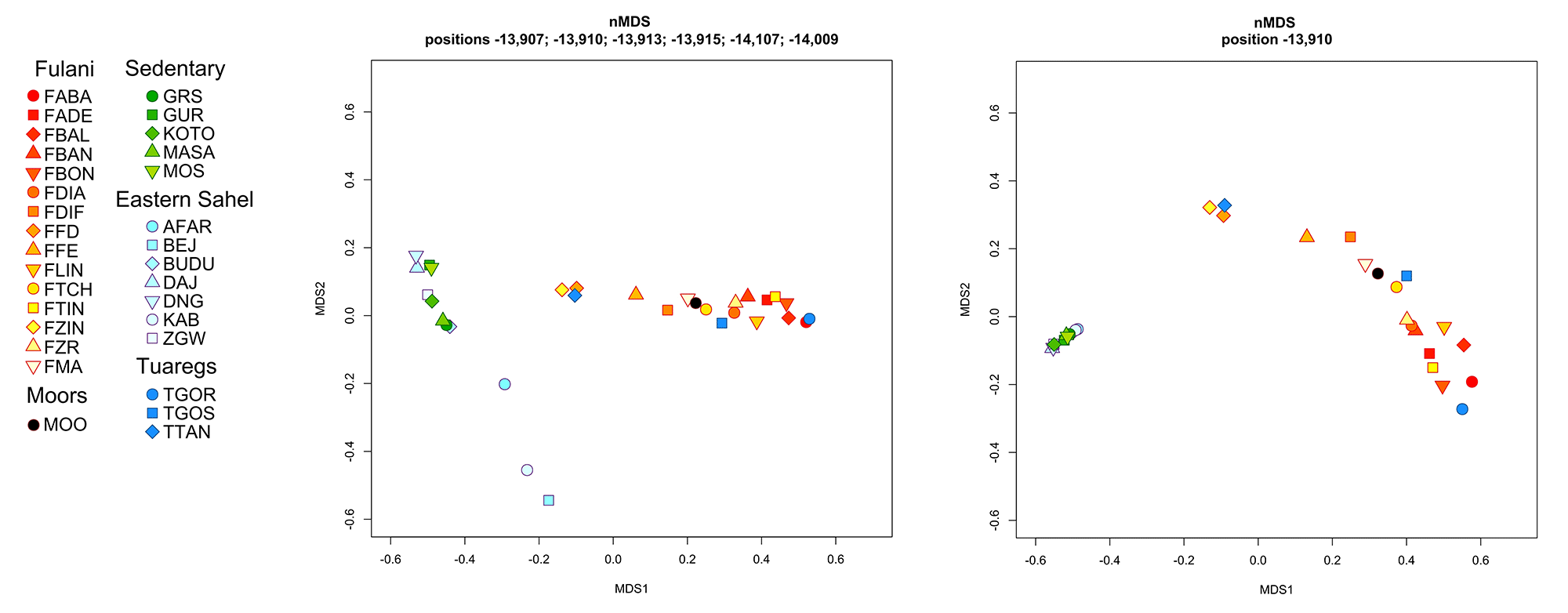
It has been suggested that the Fulani acquired the −13910*T haplotype by admixing with a North African population which had some Eurasian ancestry, so that this variant in the Fulani population has the same descent as the variant found in Eurasia (Coelho et al., 2005; Mulcare et al., 2004; Ranciaro et al., 2014). Furthermore, it has been estimated that two distinct admixture events took place, one 1,828 and the other 302 years ago (Vicente et al., 2019). Our estimates of the growth and selection of −13910*T haplotype in the Fulani indicate an earlier dating than the abovementioned admixture times (Vicente et al., 2019) and are consistent rather with the estimates of Coelho et al. (2005), who proposes a period of 10,125–6,060 years ago.
The age of the second main LP Sahelian variant −13915*G, which we estimated at 1.5 ka (Priehodová et al., 2017) perfectly matches the timing of migration of Bedouin tribes from Arabia to Africa (Zeltner, 2002). A previous estimate of the age of −13915*G, which was up to 4 ka (Enattah et al., 2008), concerns its origin in Arabia, probably Central Arabia. From there, it expanded at the beginning of Middle Ages, therefore much later than the −13910*T variant, which was already established in the western Sahelian populations.
Because the −13910*T haplotype associated with LP is shared between the Fulani from Ziniaré, Burkina Faso, and Europeans (Vicente et al., 2019) and since we found an old age of expansion for this mutation within the African population of the Fulani (dataset covering much larger area of the Sahel), it is possible that the −13910*T may have originated in a food-producing population that lived in the Near East, including northeastern Africa, as suggested by Myles et al. (2005). Subsequently, though, the first Neolithic farmers who lived in the Near East/northeastern Africa were replaced by various population movements (Černý, Čížková, Poloni, Al-Meeri, & Mulligan, 2016; Vyas et al., 2016), with the result that only the originally “Arabian” –13,915*G LP variant is present in this region today (Priehodová et al., 2014).
Interestingly, genetically the nearest population to the first Near Eastern farmers has been conserved in Sardinia (Chiang et al., 2018; Sikora et al., 2014), where the frequency of −13910*T is as low as elsewhere in the Mediterranean (Gerbault, 2013; Meloni et al., 2001). Since there is archeological evidence of milking among pastoralists in Central Sahara by at least around 7.5 ka (Dunne et al., 2012), which corresponds to our molecular estimates for the Fulani, it is possible that the first Neolithic sedentary populations consumed not fresh milk but rather dairy products such as cheese, where lactose content is low. Fresh milk consumption is, after all, a habit more common among nomadic pastoralists originating in northeastern Sahara than among sedentary farmers whose origin is in the Near East.
See also
More on the “Eurasian” LP allele:
- “Local” Tollense Valley warriors rich in haplogroups R1b and I2a
- mtDNA, lactase persistence, and admixr for ADMIXTOOLS
More on the Prehistoric Sahel and Afroasiatic:
- Visualizing phylogenetic trees of ancient DNA in a map
- Fulani from Cameroon show ancestry similar to Afroasiatic speakers from East Africa
- Sahara’s rather pale-green and discontinuous Sahelo-Sudanian steppe corridor, and the R1b – Afroasiatic connection
- Ancient genomes from North Africa evidence Neolithic migrations to the Maghreb
- Tales of Human Migration, Admixture, and Selection in Africa
- Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations
- Genetic ancestry of Hadza and Sandawe peoples reveals ancient population structure in Africa
- R1b-V88 migration through Southern Italy into Green Sahara corridor, and the Afroasiatic connection
- Potential Afroasiatic Urheimat near Lake Megachad